
In this topic, we focus on the physiology of muscle and muscular contraction and the effects of exercise. You will learn:
- muscle structure and components
- muscular contraction
- muscle fibre recruitment
- muscle fibre types
- genetics and muscle physiology
- muscular force, velocity and power
- muscular adaptation and exercise.
Terminology and vocabulary reference guide
As an allied health professional, you need to be familiar with terms associated with basic exercise principles and use the terms correctly (and confidently) with clients, your colleagues, and other allied health professionals. You will be introduced to many terms and definitions. Add any unfamiliar terms to your own vocabulary reference guide.
Activities
There are activities throughout the topic and an end of topic automated quiz. These are not part of your assessment but will provide practical experience that will help you in your work and help you prepare for your formal assessment.
Let's start with exploring the inner workings of muscle tissue, the components that make up a muscle, building up to how a muscle contraction occurs, and also how the nervous system links to muscle in order to cause contraction, hold on for the ride!
Let's begin with the function of muscles.
Functions of muscles
Muscles have a variety of roles within the human body, the main ones being:
- Producing body movements. This includes movements such as whole-body movements like walking or running and also more precise movements such as picking up a pencil or nodding your head.
- Support of internal organs: Muscles support and stabilise the spinal column and maintains posture by holding joints in place.
- Storage of substances: Storage is achieved by circular muscles called 'sphincters. For example, they prevent outflow from organs and enable temporary storage of substances in organs such as the urinary bladder and stomach.
- Protection: Sphincters of the eyes, mouth and other orifices also prevent outside foreign bodies from entering the body (for example, dust in your eyes). Muscles can also protect internal organs that don't have the benefit of bony coverings (for example, abdominals).
- Heat production: Approximately 85% of body heat is produced by muscles. As muscles contract heat is produced through a process called thermogenesis. Contrary to popular belief, this heat production is not the result of friction. When muscle contracts and uses energy, heat is produced as a by-product of cellular metabolism. When you are cold, your body will initiate this without conscious thought (that is, shivering).
In order for the muscles to perform their functions, they typically need to contract which requires innervation by the nervous system which carries impulses to the muscles. We will now look in-depth, at the process by which muscles are able to contract. The process of muscle contraction requires a few different elements/ components and a few steps which are outlined below.
Introduction to muscle contraction: Part 1
Motor neurons carry impulses to muscles. In this section, we will look at the connection between the motor neuron and muscle fibres.
A motor unit
Muscle fibres never fire in isolation. Instead, a group of fibres is innervated by a single motor neuron.
A motor unit is defined as one motor neuron and all the muscle fibres it innervates.
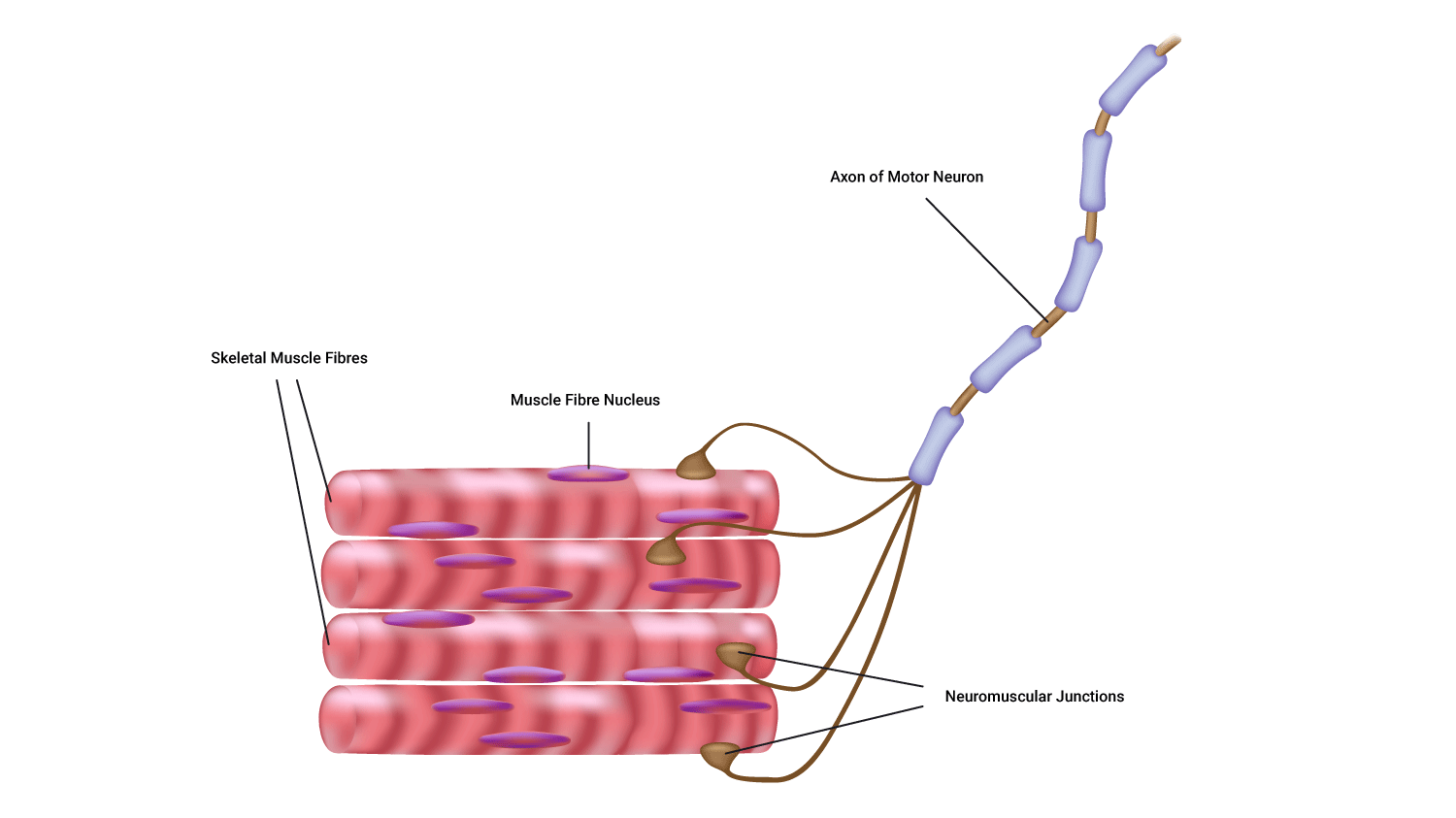
Parts of a motor unit
- One motor neuron
- Muscle fibres
- The neuromuscular junctions.
Structure of a synapse
A synapse is the site of communication between a neurone and another neurone or a neurone and an effector cell, allowing communication.
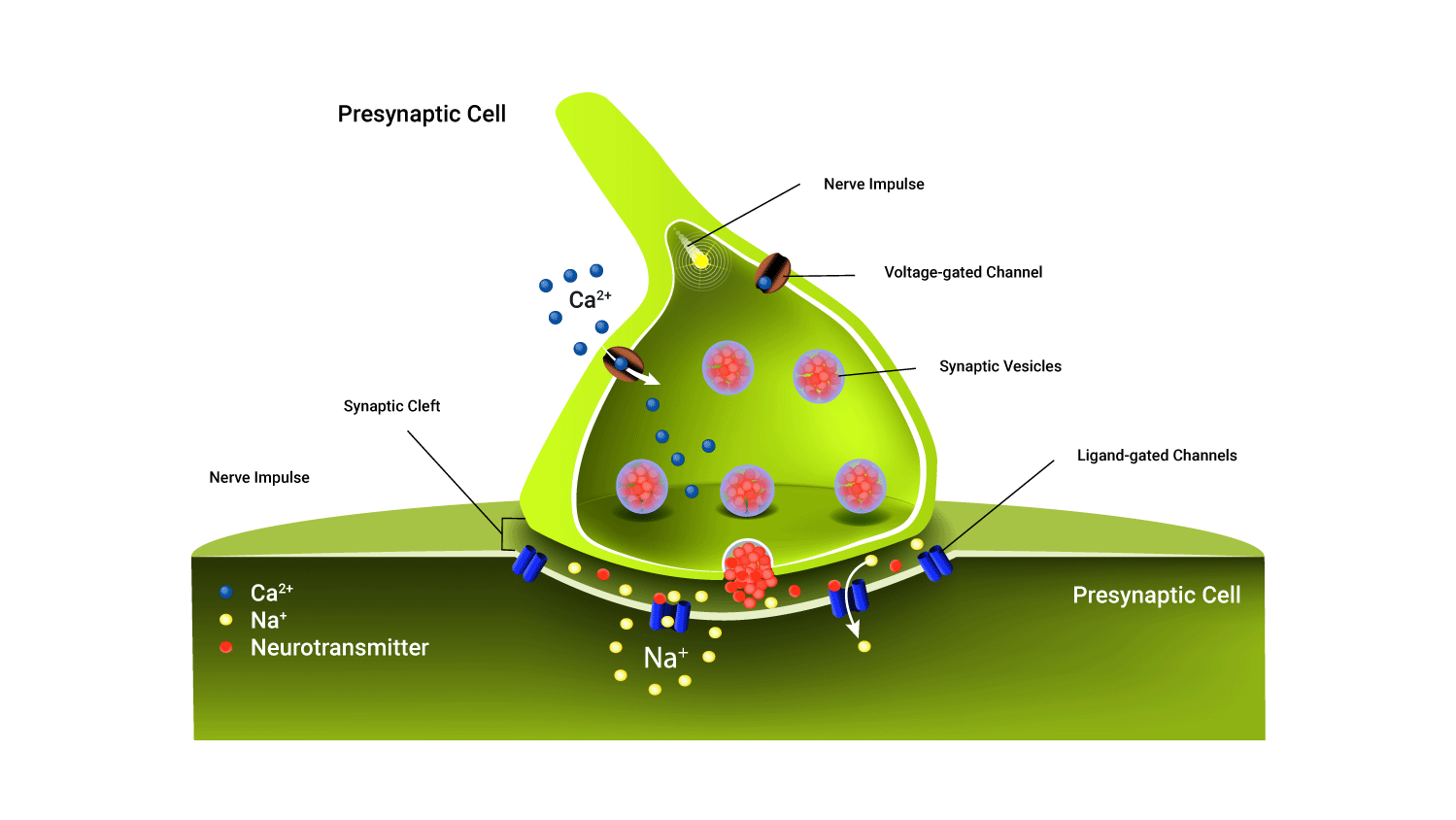
These meeting points allow impulses to be passed between the different neurons. Transmitting these electrical impulses contain a few steps and are somewhat complex at first, but we will break each of the steps involved below.
- The pre-synaptic terminal: This is the synaptic end bulb (or knob) of the first neuron. The term 'pre-synaptic' describes its location as coming before the synapse. It contains neurotransmitters packaged into 'vesicles'. Neurotransmitters are chemical couriers that are vital in generating new impulses on the other side of the synaptic cleft.
- The synaptic cleft: This is a small gap that separates the pre and postsynaptic terminals (that is, the connecting parts of two neurons).
- Impulses can't 'jump' the synaptic cleft, instead, they send neurotransmitters across.
- The postsynaptic membrane: The dendrite of the next neuron, (post~ after). When neurotransmitters bind to receptors on the postsynaptic membrane, ion channels open allowing ions to enter (typically positively charged ions). If enough arrive a new impulse is formed and the message is passed on.
In the case of the motor neuron meeting a muscle, the post-synaptic membrane is the muscle fibre. Instead of being known as a synapse, this meeting point is known as a neuromuscular junction, which we will now discuss.
Neuromuscular junction
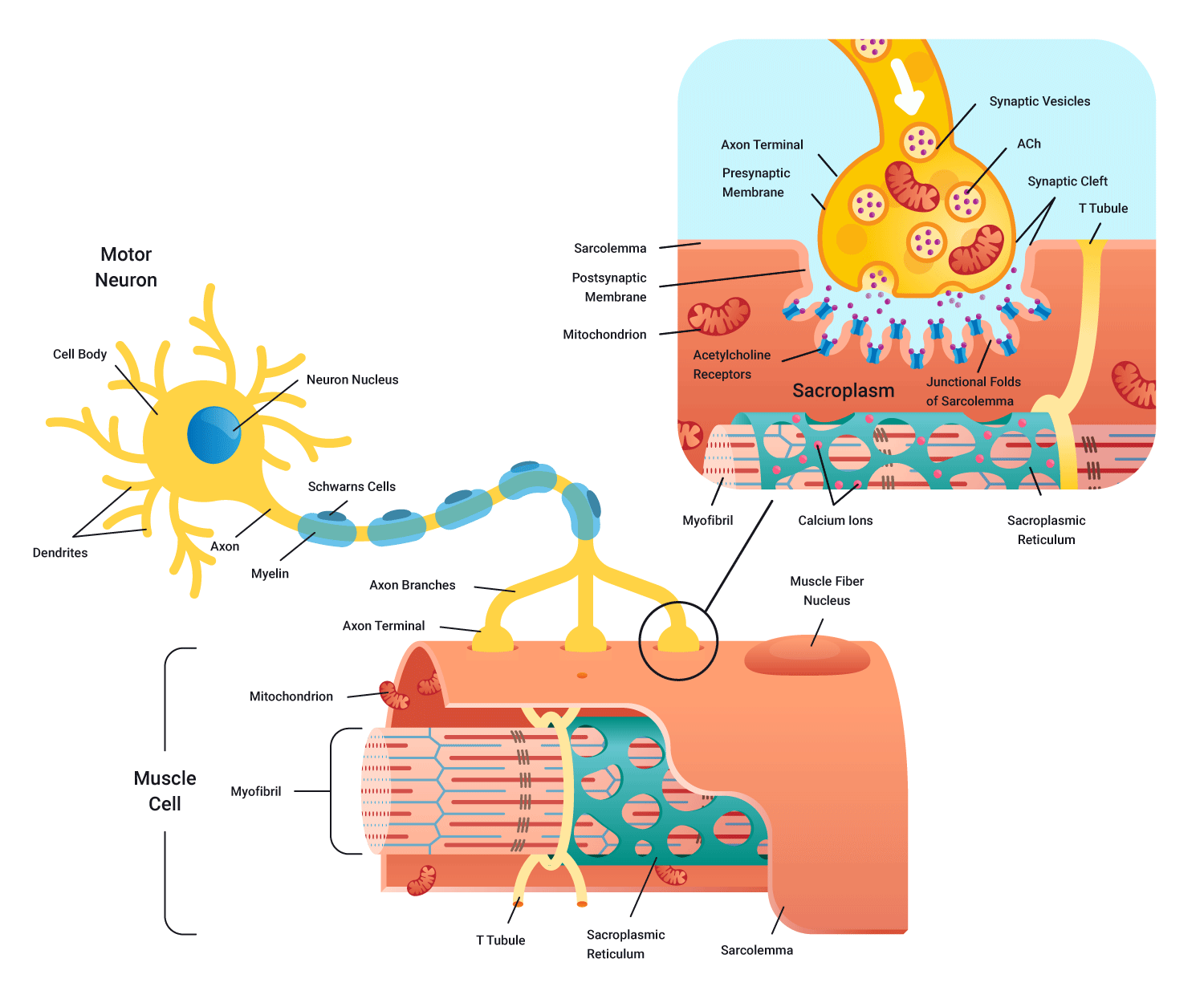
As you can see in the preceding diagramme, the synaptic cleft (gap) still exists in this meeting of nerve and muscle. As impulses (action potentials) cannot jump the gap, we require a neurotransmitter to cross the cleft in order to stimulate a charge on the other side. In this case, the neurotransmitter is known as acetylcholine (Ach).
As in the case of a message being passed between neurons (at the muscle), motor neurons arriving from the spinal cord arrive at the pre-synaptic terminal causing the release of the neurotransmitter acetylcholine (Ach) across the neuromuscular junction. Acetylcholine then binds to receptors on the muscle fibre surface.
A new electrical charge is then transmitted through the full length of the muscle fibres in that motor unit. These impulses could potentially lead to contraction.
We will now do an overview of a neurone to muscle communication (pre-contraction). Think of this as Part 1 of muscle contraction with Part 2 being the Sliding filament theory.
How nerves fire (Muscle contraction: Part 1)
Electricity (stimulus) is the main currency required for muscle contraction to occur, along with sodium(Na+) Calcium (Ca2+), ATP ( Adeonsidetriphosphate- energy currency), and the neurotransmitter acetylcholine (Ach). Let's look at the main steps.
- Electrical impulse (action potential) flows down the body of the presynaptic neuron
- This positive charge (voltage) encourages ion channels on the membrane of the synaptic end bulb to open and let in calcium Ca2+.
- Ca2+ triggers the packages (vesicles) of neurotransmitters to move from the middle of the bulb to the outer membrane and spill their contents into the synaptic gap. Note: Sodium Na+ is already 'floating' in the gap.
- The neurotransmitters then bind to receptors on the postsynaptic terminal (bulb) and trigger the receptors on the postsynaptic membrane to open.
- Sodium ions (Na+) which were floating around in the synaptic gap can now flow into the postsynaptic membrane and flood the membrane with a positive charge. Note: This positive charge needs to be of a sufficient level in order to 'depolarise' the membrane (that is, to stimulate the membrane)...To be continued in Part 2.
Let’s pause for a second and take some time to familiarise ourselves with the physiology and the microscopic structures of a skeletal muscle fibre, we will continue Part 2 of muscle contraction when we look more in-depth at the sliding filament theory.
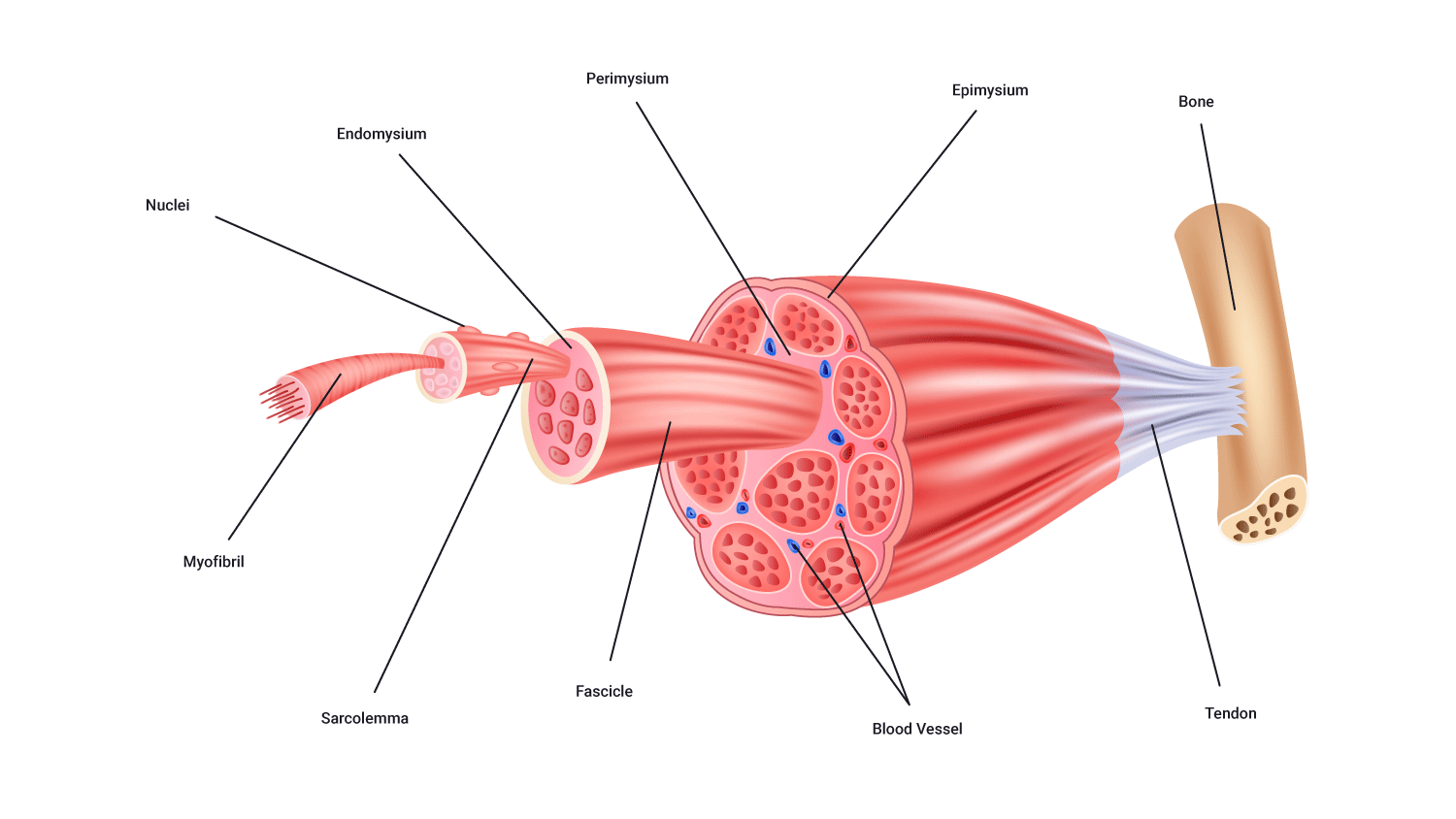
Muscle structure: The main players
Let's now focus on the individual structures that collectively make up a muscle. It is important to note that many of these structures are smaller than what we can pick up with the naked eye. For you to get a complete understanding of muscle structure we will start from the outside and work our way in.
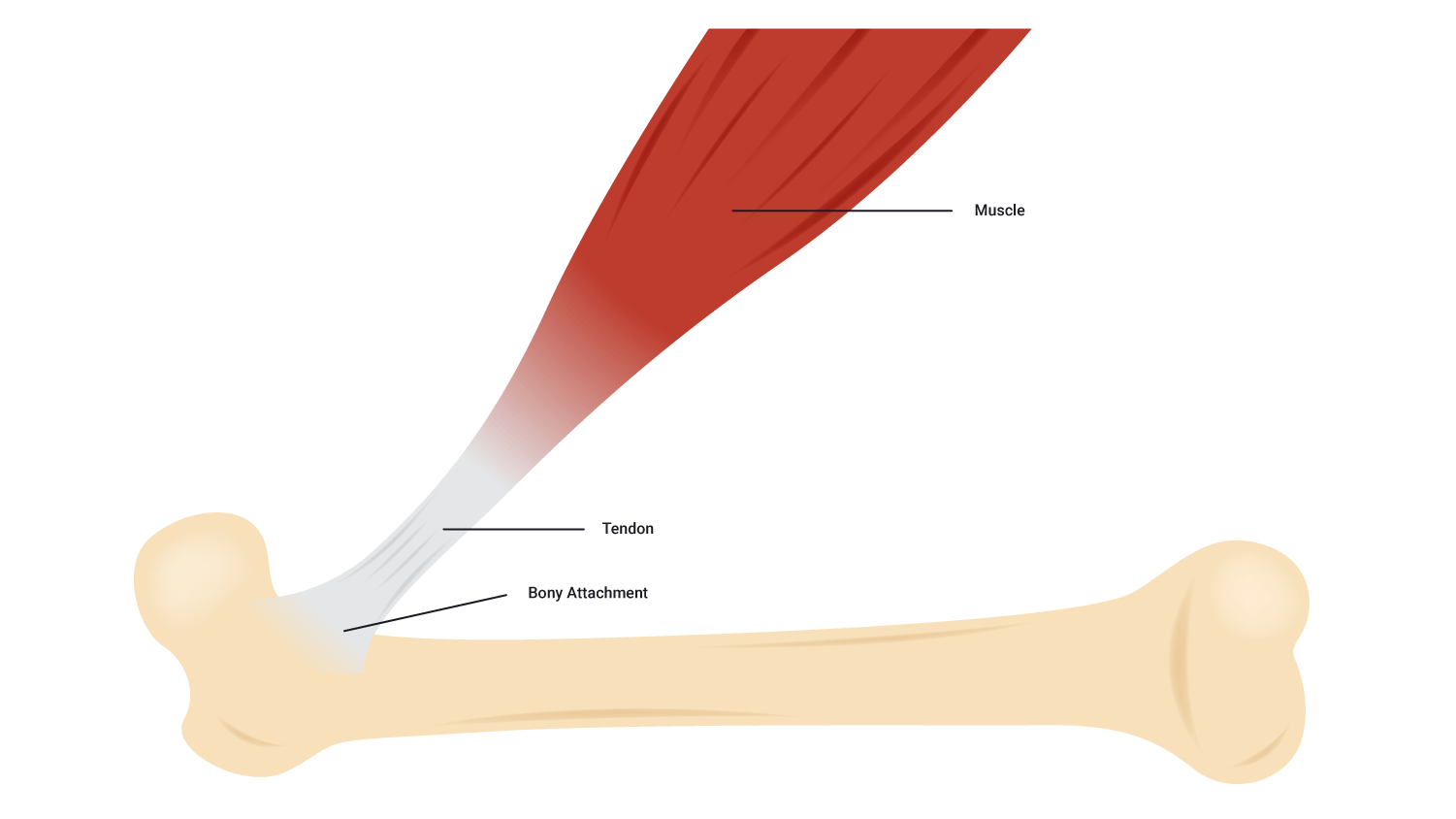
Tendons
- Tendons are tough cords of collagen fibres that grow directly from the muscle fibres which attach muscle to bone.
- Without this link, there would be no limb movement.
Fascia
- The fascia is a protective connective tissue that surrounds muscles, bones, organs, nerves, blood vessels, and other structures. It also separates the various tissues from one another.
- The facia holds muscles with similar functions together. It allows 'free' movement' of muscles. That is, it carries structures such as nerves, blood vessel and lymphatic vessels as well as filling the spaces between muscles.
- It is also responsible for maintaining structural integrity; for providing support and protection; and acts as a shock absorber.
Three layers of connective tissue extend from the facia, which are named based on their location:
- Epimysium (Epi~ on/upon/before): This is the outer layer of connective tissue surrounding the whole muscle.
- Perimysium (Peri~ around/enclosing): This internal layer surrounds groups of 10-100 muscle fibres, bundling these fibres up into clusters or bundles called fascicles.
- Endomysium (Endo~ within). This layer surrounds each individual muscle fibre and separates each one from another.

Muscle fibres
- Muscles are made up of a large number of individual structures called fibres.
- The number of fibres present in a muscle varies depending on the size and function of the muscle.
- Muscle fibres range in diameter from 10 to 80 micrometers which are practically invisible to the naked eye. However, they can be more than 35 cm long.
Fascicles
- Muscle fibres are known as myocytes (Myo~=muscle).
- Within a muscle, they are organised into bundles known as fascicles.
- Each fascicle is likely to be made up of one type of muscle fibre (for example, slow twitch and fast twitch).
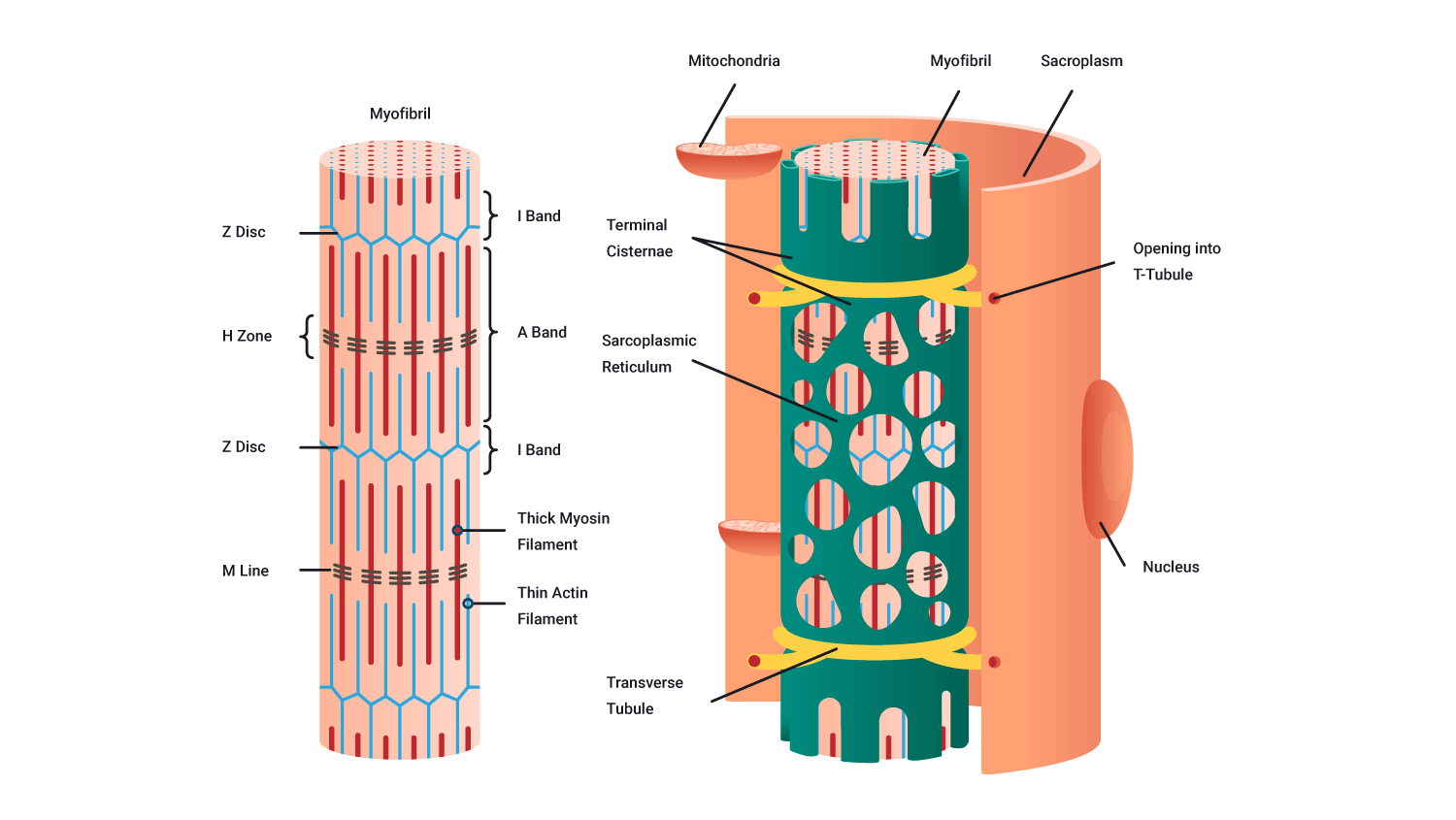
Sarcolemma
- A muscle fibre surrounded by a plasma membrane is called the sarcolemma.
- Inside the sarcolemma, there are myofibrils which are rod-like structures running the length of the muscle fibre.
- There is also a gelatine-like substance that fills the spaces between the myofibrils. This is the sarcoplasm. This is where fuels are stored, such as glycogen, amino acids, and triglycerides. You can see these in the following diagramme.

Sarcoplasmic reticulum
- The sarcoplasmic reticulum is a longitudinal network of tubules (green in the following picture).
- These lie parallel to the myofibrils and loop around them.
- It is a storage site for calcium (Ca2+) which is essential for muscle contraction.
Myofibrils
- Each muscle fibre consists of several hundred to several thousand myofibrils.
- Myofibrils are the contractile elements of skeletal muscle.
- They appear as long strands of even smaller sub-units called sarcomeres.
Sarcomere
- Myofibrils are broken up along their lengths into sarcomeres.
- It is the interaction between the two myofilaments found within these repeating sub-units that we need to understand in order to fully appreciate how a muscle contracts.
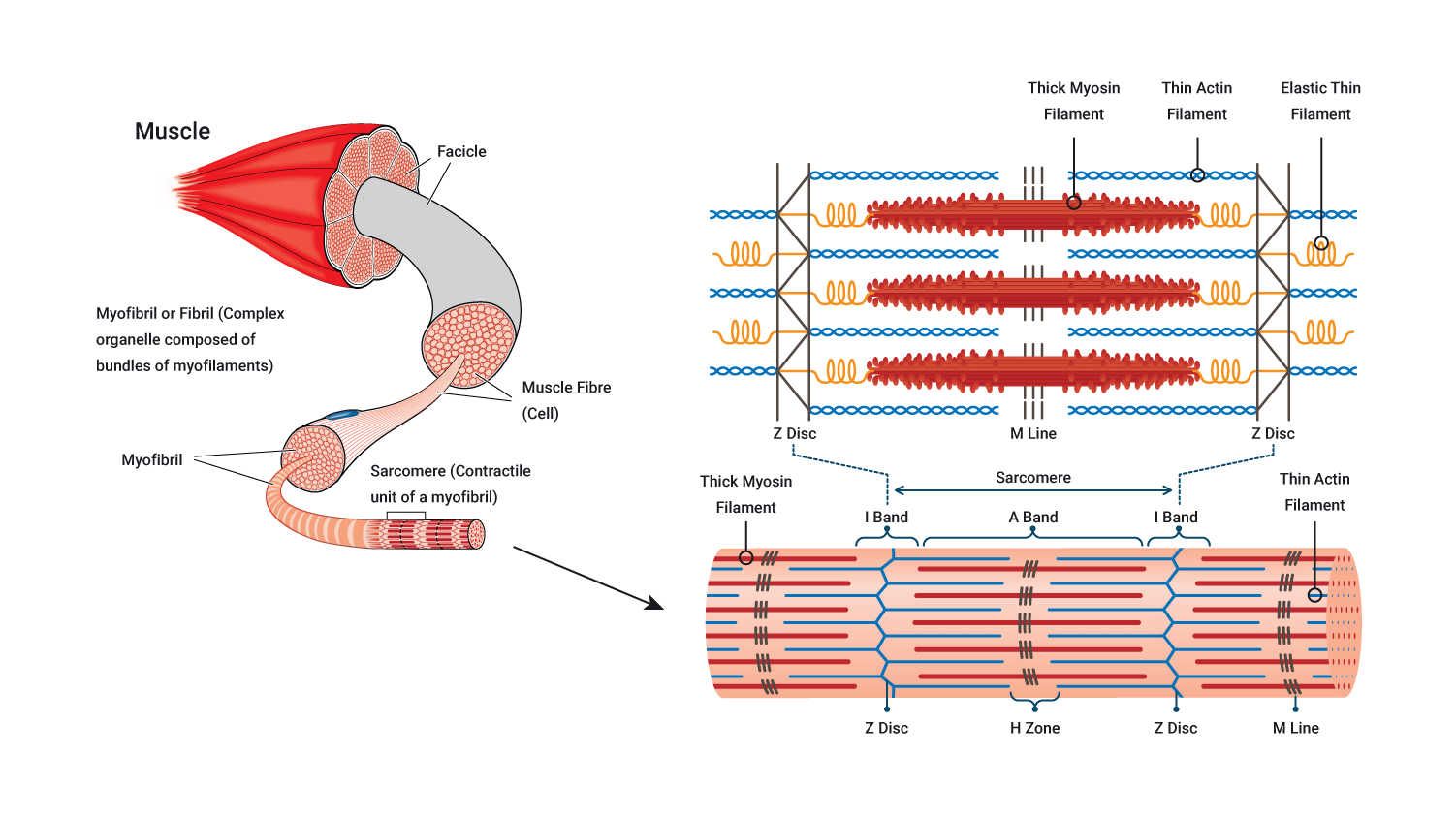
Myofilaments
Myofibrils are composed of two types of myofilament called myosin (also known as contractile proteins) and actin:
- Myosin is a thick protein filament.
- Actin is a thin protein filament.
It is these myofilaments that interact to cause contraction of a sarcomere, which translates to contraction of a myofibril, muscle fibre and in the end the muscle itself. Let's look further at these.
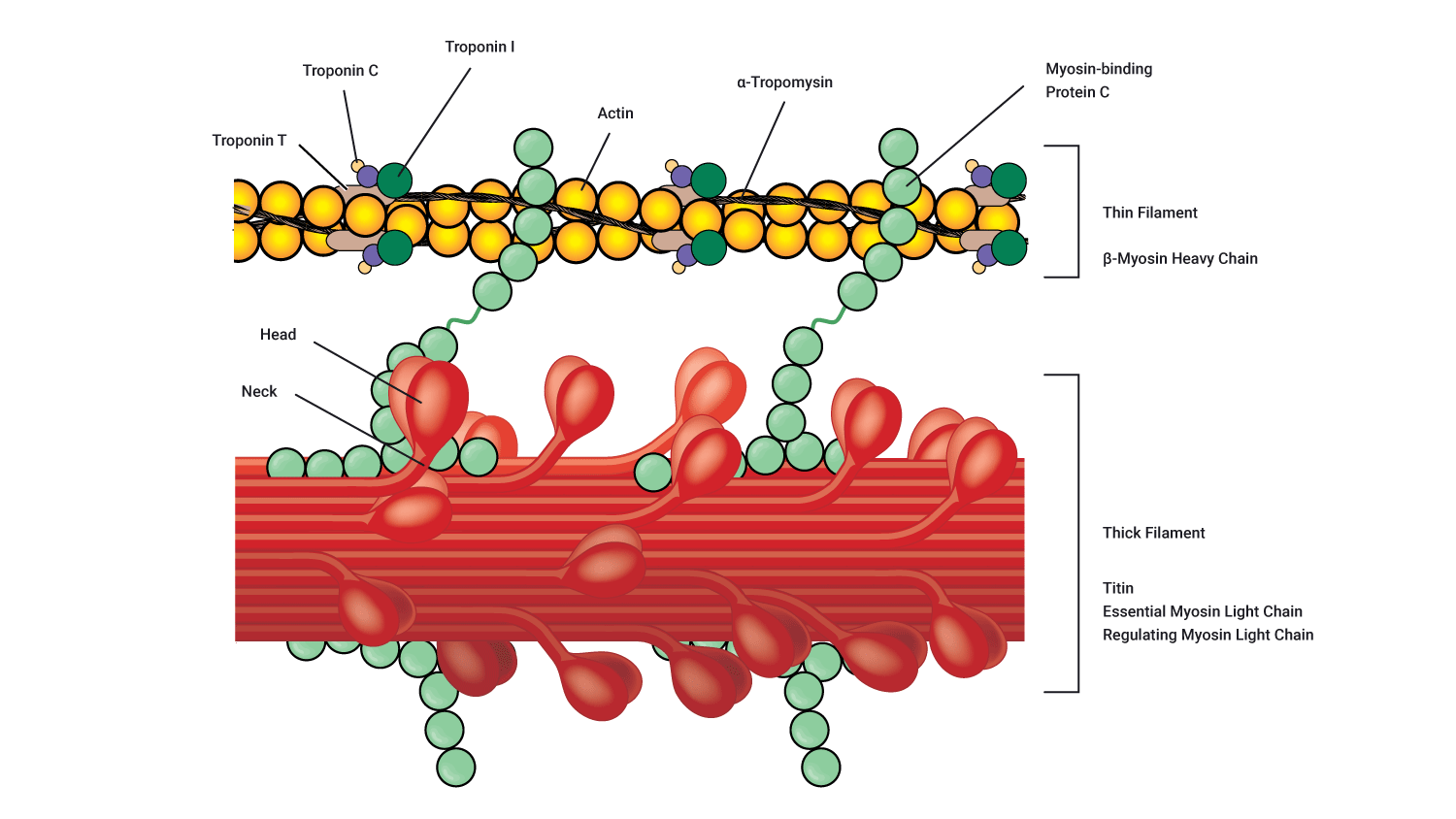
Myosin
Each myosin filament contains several hundred globular heads, which protrude from the myosin filament to form cross-bridges that interact during muscle contraction with special active sites on the actin filaments. These globular myosin heads are shown in purple on the following diagramme.
Actin
Each actin filament contains active sites to which the myosin head can bind. At rest, these active binding sites are covered by a barrier known as tropomyosin. Tropomyosin is guarded by a molecule known as troponin.
Muscle contraction: Part 2
The process of muscle contraction is described by a theory called the 'Sliding filament theory'. The theory suggests that the two myofilaments actin and myosin slide past each other during muscle contraction. Before we get into the nitty-gritty of the process, let’s take a look at all the players in the game.
- Calcium: Calcium is one of two substances vital for muscle contraction. Calcium binds to troponin which stimulates tropomyosin to move and reveal to the myosin-binding sites on actin.
- ATP: The second of two energy sources required for muscle contraction. Adenosine triphosphate (ATP) binds to the myosin head and provides the energy required for myosin to move, bind to actin and perform the 'power stroke'.
- Troponin and tropomyosin: Collectively known as the regulatory proteins because, in a resting state, they cover the myosin-binding sites on actin and act as the regulator for muscle contraction. In order to contract a muscle, myosin heads need to connect to actin. In a resting state, tropomyosin covers the binding sites on the actin filaments preventing the binding of myosin heads.
- Actin and myosin: Collectively known as the contractile proteins because when stimulated, these components are responsible for muscle contraction by binding together and 'sliding' over each other (hence the term, 'Sliding filament theory').
- Sarcoplasmic reticulum: Stores a high volume of calcium (Ca2+) when a muscle is at rest. When stimulated, the sarcoplasmic reticulum releases its calcium which enables the remainder of the steps required for the contraction to continue.
The Sliding filament theory: Behind the scenes of the process
Let's review and build on what we have learnt so far...
- The process begins with the arrival of the motor impulses at the neuromuscular junction.
- The neurotransmitter acetylcholine (Ach) is released across the synaptic cleft which binds to receptors (channels) on the postsynaptic membrane.
- These channels then open and permit the entry of sodium (Na+) into the postsynaptic membrane. The arrival of sodium (Na+) increases the positive charge within this region and as a result, stimulates an action potential.
- This action potential travels throughout the portions of the muscle and heads towards the sarcoplasmic reticulum (huge calcium storage found wrapped around the myofibrils within a muscle fibre) which becomes stimulated by these new impulses.
- The result of this is a release of calcium (Ca2+) into the myofibrils of the muscle. The contraction process continues via the involvement of the:
- contractile proteins actin and myosin
- regulatory proteins troponin and tropomyosin
- energy source known as ATP.
- Calcium stimulates the troponin to trigger tropomyosin to move (tropomyosin covers the myosin-binding site when a muscle is not contracting) and by doing so, reveals the myosin-binding site on the actin.
- ATP then binds to the myosin heads which gives them the energy they need to move, bind to their site on actin allowing myosin heads to form cross-bridges with actin and start the contraction process.
- When myosin forms cross-bridges with actin, the two filaments slide past one another as actin is pulled towards the centre of the sarcomere. It is thought that the myosin heads (cross-bridges) literally pull the actin filaments towards the centre of the sarcomere, therefore shortening the muscle. The tilting of the myosin head is called the power stroke.
The following diagramme presents this information visually.
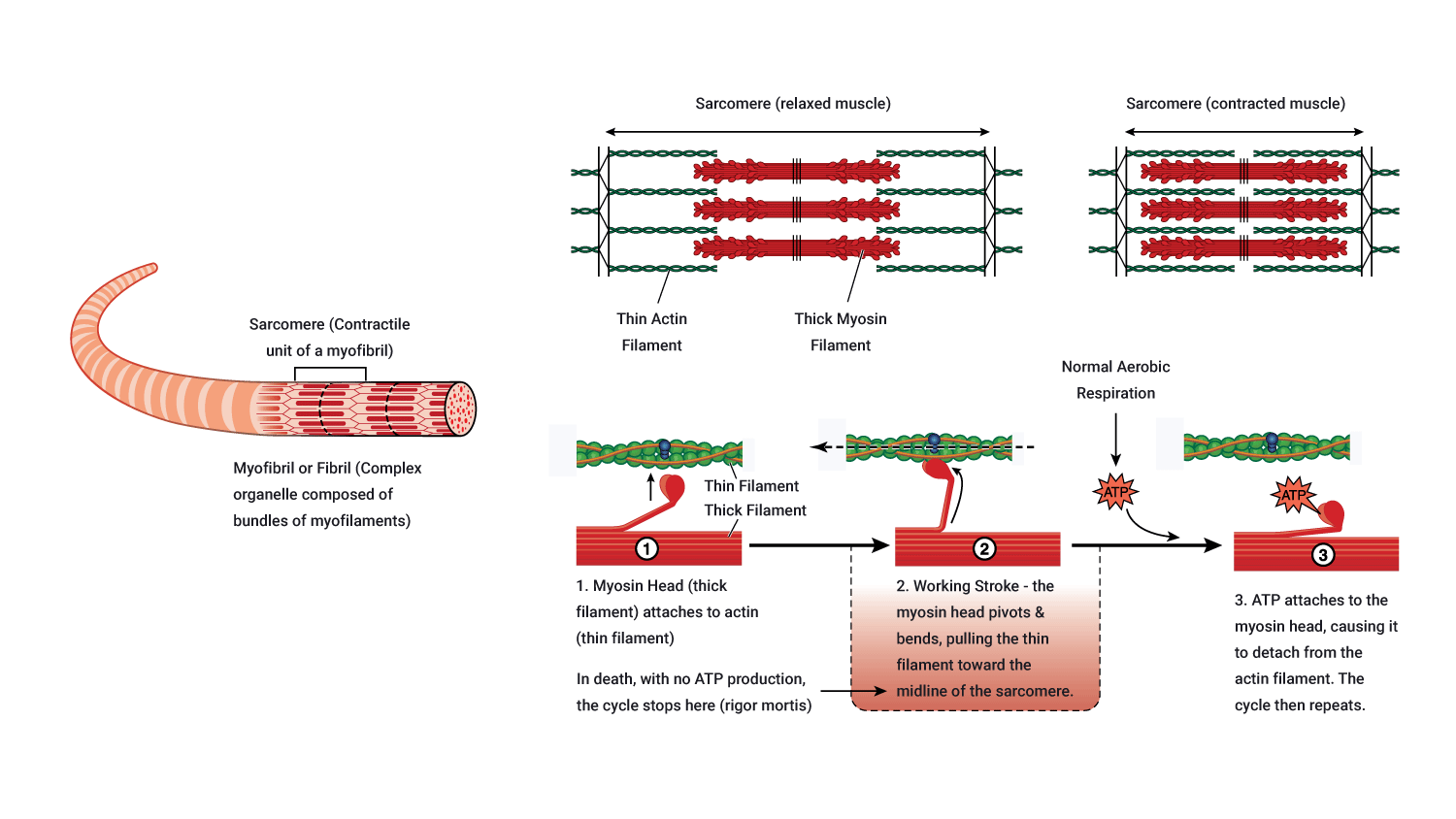
Muscle shortening
Immediately after the power stroke, the myosin head breaks away from the active site, rotates back into its original position and attaches to a new site further along the actin filament.
Repeated attachments, power strokes and re-attachments cause the filament to slide past one another (hence, the Sliding filament theory!) and muscle shortening occurs. Around three to five (3-5) power strokes are performed by each myosin head per second of contraction! Crazy!
Ending a muscle contraction: Key steps required for muscle relaxation
A muscle will continue to contract all the time there are sufficient levels of Ca2+, ATP and electrical impulses (action potentials) arriving. After the last action potential arrives, with the support of ATP, calcium (Ca2+) is pumped back into the sarcoplasmic reticulum where it is stored in preparation for the arrival of the next action potential. As this requires energy in the form of ATP. It is worth noting that ATP is needed for both muscle contraction and relaxation.
During the phase of muscle relaxation, tropomyosin moves back into its 'resting position' covering the myosin-binding sites on actin, preventing any cross-bridge forming. Until the next action potential, there are insufficient volumes of calcium (Ca2+) and ATP, therefore muscle contraction is unable to occur.
Steps required to aid muscle relaxation
- The enzyme acetylcholinesterase breaks down and decomposes acetylcholine. This neurotransmitter can no longer stimulate the muscle in a decomposed state.
- Any remaining or residual calcium ( Ca2+) is pumped back into the sarcoplasmic reticulum (with the aid of ATP).
- Troponin and tropomyosin return to their original positions, re-covering the myosin-binding sites on actin.
- ATP is broken down into its constituents which results in the disconnection of the cross-bridge connection between actin and myosin to break down (no longer connecting these two contractile proteins).
- Muscle fibre relaxes.
Summary of muscle contraction
The following video, whilst slightly long, simplifies this process and provides entertaining analogies to support our understanding of this process. Again, you do not need to know this in-depth but it is important that you have a grasp on the process overall. Enjoy the video below, test yourself by writing down the steps involved and seeing if you can place them in the right order.
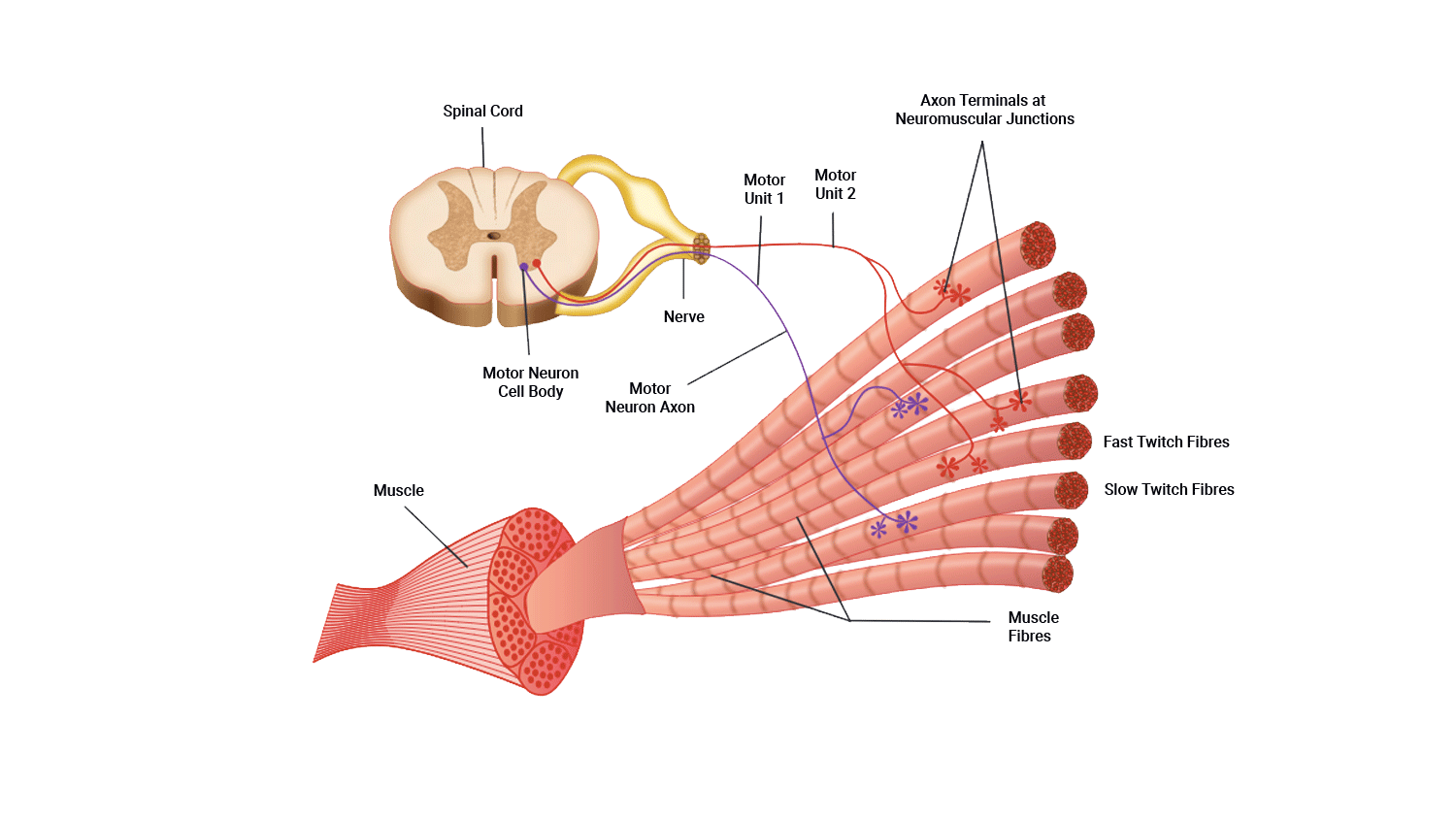
This section describes how muscle fibres are recruited.
Motor unit recruitment
Now that we understand how muscle fibre contracts, we need to broaden our focus as single muscle fibres never contract alone.
Muscles are made up of fascicles full of fibres. One motor unit can fire multiple fascicles of fibres if they belong to the same motor unit. If the impulses carried by the motor unit reach the muscle fibre threshold then all muscle fibres attached to that motor neuron will contract 100%.
The number of muscle fibres in a given motor unit depends on the size of the muscle and the muscles location and function. For example, in the small and large muscles.
Small muscle
One nerve fibre innervates a few muscle fibres. For example, in an eye muscle, one motor neuron may innervate 23 muscle fibres.
Large muscle
One nerve fibre innervates more muscle fibres. For example, in quadriceps, one motor neuron may innervate 500 muscle fibres.
The 'all or nothing' principle
The 'all or nothing' principle states that the strength of a muscle fibre (or nerve cell) response is not dependant on the strength of the stimulus; this means, as long as a stimulus is above a certain stimulus, the muscle will contract.
A motor unit will contract with maximum force if the stimulus is equal to, or above, the threshold
Typically, the different motor units of a muscle fibre are not recruited all at once or at the same time. While some are contracting, others are relaxed, which delays muscle fatigue and allows for whole muscle contraction to be sustained over a longer period of time. There is a method behind nature's madness. In this model, the weakest motor units are recruited first with the stronger motor units being called upon as needed if the task requires more force.
Different motor unit types have different excitatory thresholds. This is where muscle fibre types come into play, (for example, slow-twitch vs fast-twitch fibres). Have a look at the following graph.
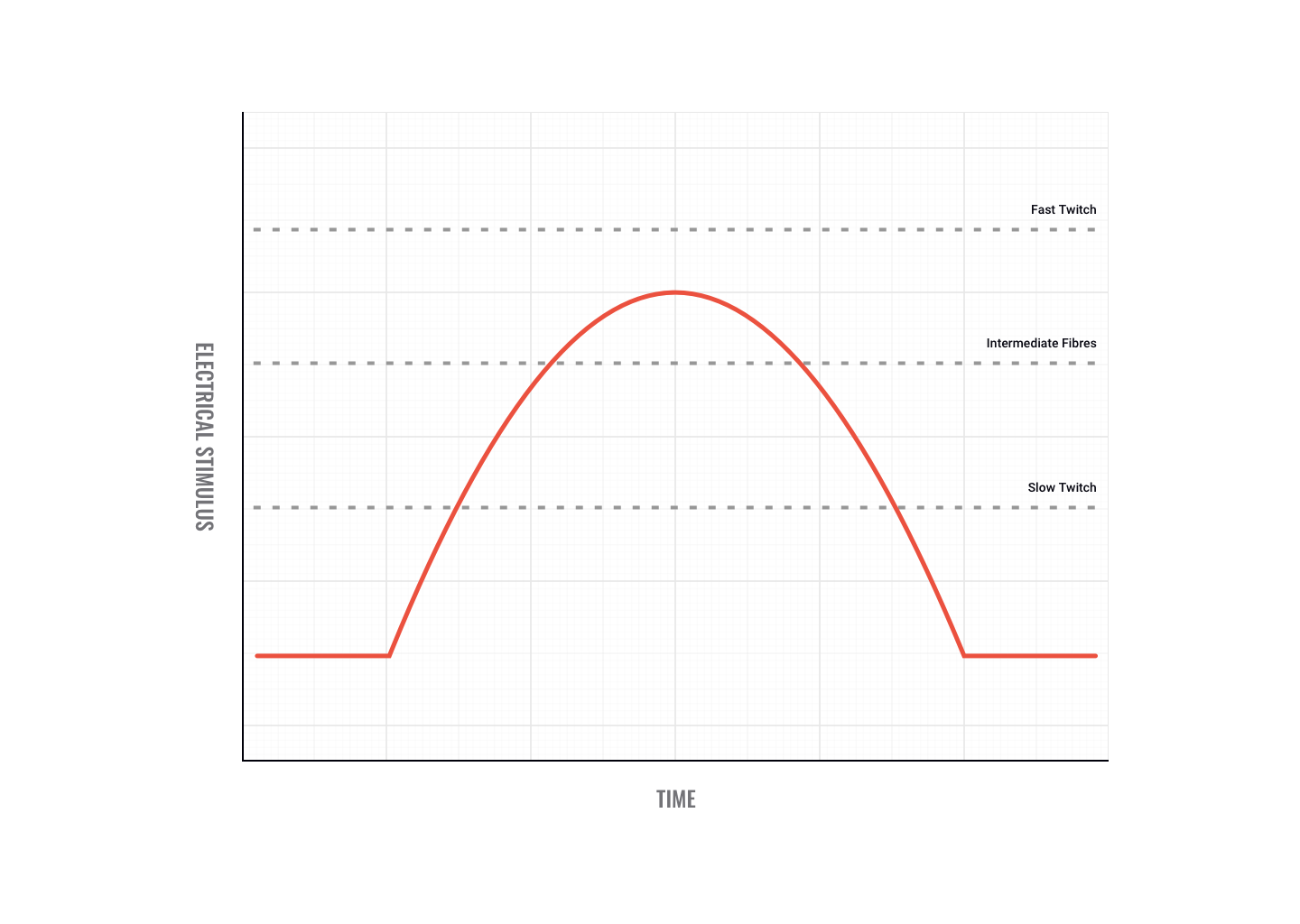
As you can see in the preceding graph, fast-twitch fibres have a higher threshold than intermediate and slow-twitch fibres. That means it takes more electricity to fire them. If the impulses sent down motor neurons to the muscle reached the threshold depicted in the graph, then only the slow-twitch and intermediate fibres would fire. It is important to note that slow-twitch fibres are innervated by slow-twitch motor neurons and fast-twitch fibres are innervated by fast-twitch motor neurons.
So if muscle fibres can only contract 100%, how can contraction strength vary?
Contraction strength
Contraction strength does not depend on how strongly individual motor units contract but how many motor units contract. When little force is needed, only a few motor units are stimulated to act. This indicates the principle of orderly recruitment.
Slow-twitch motor neurons are smaller and innervate fewer fibres than fast-twitch motor neurons. Therefore, for low-intensity activity, most muscle force is generated by slow-twitch fibres. As intensity increases, intermediate fibres are recruited and at higher intensities, fast-twitch fibres are activated. The same pattern of recruitment occurs during events of long duration.
For example, if weak contractions are sufficient to perform a task, slow oxidative fibres (SO) are recruited. If more force is needed, fast oxidative-glycolytic fibres (FOG) are called upon. If maximal force is required all fibres are called up (that is, slow oxidative, fast oxidative-glycolytic and fast glycolytic fibres (FG). Therefore, we can see, activating more muscle fibres produces more force. The nervous system recruits the number of motor units it deems necessary to create the desired response.

Muscles are made up of three main fibre types that have different characteristics.
- Slow-twitch (Type I)
- Fast-twitch B (Type IIb)
- Fast-twitch A (Type IIa) intermediate.
A couple of key terms
Mitochondria
Mitochondria are found within the muscle cell and referred to as the 'powerhouse' of a cell. The function of mitochondria is to produce ATP for muscle energy using the aerobic system.
Myoglobin
Myoglobin is a red pigment which acts as a storage site for oxygen within the muscle. Skeletal muscle fibres which have a high myoglobin content are referred to as 'red muscle fibres' and appear darker (think of the colour of chicken meat in the leg versus the wing). Wings perform a more rapid movement (that is, flight) compared to the legs which require more oxygen. As we know, structure mirrors function, therefore there is more myoglobin in the muscle fibres of the wing than of the leg resulting in a darker appearance.
Characteristics of the different muscle fibre types
Let's now look at the characteristics of the following different muscle fibre types:
- slow-twitch fibres
- fast-twitch fibres.
Slow-twitch fibres
Slow (oxidative) fibres (SO) are generally used for posture and low-intensity movement. For example, the soleus and postural muscles of the back
Intermediate fibres (fast oxidative glycolytic)
Type 2(a) fibres: Fast oxidative-glycolytic fibres (FOG) contain characteristics of both type I and Type IIb fibres. Rather than used for specific tasks, these fibres can be trained to take on more of the characteristics of one of the other fibre types.
Fast-twitch fibres
Type 2(b) fibres: Fast glycolytic fibres (FG): For specialised use in 'quick energy' activities. For example, the gastrocnemius muscle (calf) of a 100 m sprinter would have a high proportion of type 2b fibres.
The difference in muscle fibre characteristics can also have a an effect on the way a muscle presents in terms of colour.
The characteristics of the three muscle fibre types are shown in the following table.
| Slow Oxidative Fibres (SO) -Type I | Fast Oxidative-Glycolytic Fibres (FOG)- Type II(a) | Fast Glycolytic Fibres (FG)-Type II(b) | |
|---|---|---|---|
| Structural characteristics | |||
| Myoglobin content | Large amount | Large amount | Small amount |
| Mitochondria | Many | Many | Few |
| Capillaries | Many | Many | Few |
| Colour | Red | Red-pink | While (pale) |
| Motor unit size | Small | Medium | Very large |
| Functional characteristics | |||
| Capacity for ATP generation | High | Intermediate | Low |
| Contraction speed | Slow | Fast | Very Fast |
| Fatigue resistance | High | Intermediate | Low |
| Order of recruitment | First | Second | Third |
| Location where fibres are found | Postural muscles of the back and neck | Lower limb muscles | Upper limb muscles |
| Primary function of muscles | Maintaining posture & aerobic (with O2) endurance activities | Walking, sprinting | Rapid, intense movements of a short duration |
| Maximum duration of use | Hours | <30 mins | <1 minute |
| Activity used for | Aerobic | Long-term Aerobic | Short-term aerobic |
Chicken vs beef
With the information we have covered so far, it is a good time to pause and simplify things a little. Consider the following question.
Have you ever thought why some cuts of meat are dark and some cuts are lighter? For example, chicken vs beef. Think about what you now know about the characteristics of the different fibre types.
The answer to this question revolves around the amount of blood and myoglobin in a muscle. Slow-twitch fibres have more capillaries (and therefore blood) and higher levels of myoglobin (a red pigment). Therefore, muscles composed of slow-twitch fibres are red in nature. Fast-twitch fibres have fewer capillaries and less myoglobin. Therefore muscles made up of mainly fast-twitch fibres are lighter in colour.

Now consider for a moment what a cow does all day? It stands and eats. Cows are predominantly slow-twitch orientated animals (have you ever seen a cow jump or sprint?). Beef is very red in nature as it is full of blood and myoglobin.
Chicken: light and dark meat
Chicken is an interesting example. Breast meat is very light in colour, whereas leg meat is darker. This makes sense when you think about how these areas are used. Breast meat is obviously attached to wings, which chickens use to quickly fly away when spooked (fast-twitch orientated), whereas legs are for walking and roosting (slow-twitch orientated) and therefore have more blood within.

Humans
Humans also have darker and lighter meat. Some of our muscles, like the soleus in the lower leg are almost all slow-twitch fibres. Others such as those controlling eye movements are made up of only fast-twitch fibres. Muscles adapt to the function they are required for. The majority of human muscles contain a mixture of both slow and fast fibre types.

Fibre type allocation
Essentially, your fibre type allocation is dependent on your genetics. From an evolutionary standpoint, this makes sense. Our not so very distant ancestors daily survival sometimes dictated a long walk or jog in search of food. Other times, a fast sprint or jump may have kept one out of harm's way. The exact composition of each muscle is genetically determined.

Africans and descendants of Africans have dominated world athletics events at both ends of the spectrum. Marathon running has been dominated in recent years by African countries. Sprinting has been dominated in recent years by descendants of Africans now living in the Caribbean, the USA and Britain. Genetics has a part to play in this.


Unfortunately, science suggests you cannot significantly alter your fibre allocation. You may have to thank your parents for your sporting success or failure.
If you want to win an Olympic medal in the 100 metres, you had better be born with about 80% fast-twitch fibres in the key muscles! Want to win the Olympic marathon? Put in an order for 80% slow-twitch fibres in your quads.
Let's start with a recap.
Recap
Individual muscles are a mixture of three types of muscle fibres (type 1, type 2a and 2b), but their proportions vary depending on the location and action of that muscle.
It must be remembered that skeletal muscles, although a mixture, can only have one type of muscle fibre within a motor unit. This is demonstrated if we look at contractions.
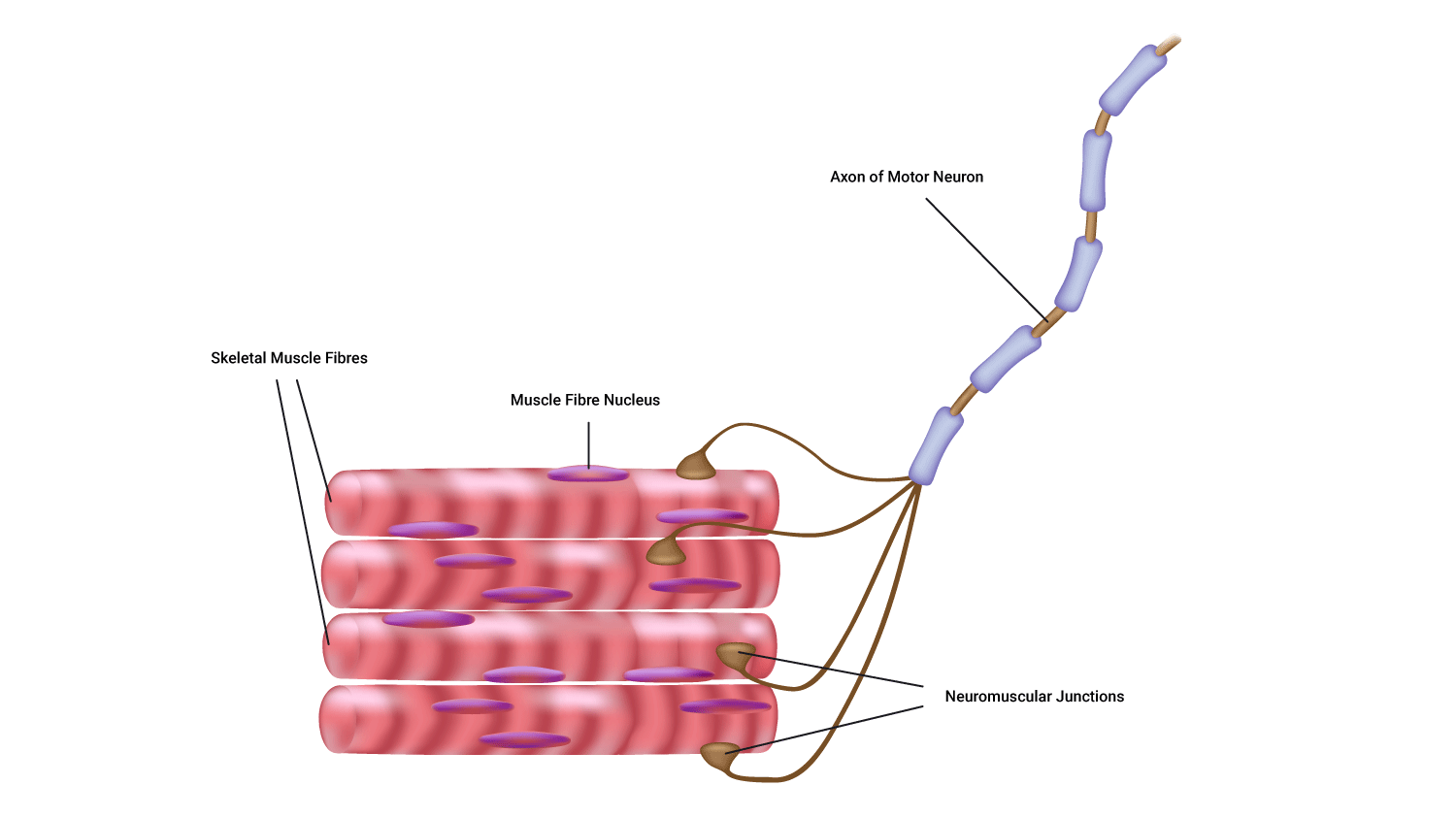
Muscle fibre recruitment
- If a weak contraction is needed only the type 1 motor units will be activated. These fibres are used mainly for endurance activities.
- If a stronger contraction is required the type 2a fibres will be activated or used to assist the type 1 fibres.
- Maximal contractions facilitate the use of type 2b fibres which are always activated last. These fibres are used during ballistic or strength activities but tire easily.
Can you convert fibres?
In short, no. Traditionally, the total number of skeletal muscle fibres has been thought not to change.
While intermediate fibres will act like either slow-twitch or fast-twitch depending on the type of training undertaken, they do not actually convert. Similarly, slow-twitch fibres can not become fast-twitch and vice-versa, although they can develop some characteristics of other fibre types (for example, fast-twitch fibres can develop more resistance to fatigue if long term endurance training is undertaken).
The main reason why fibres do not convert appears to be mainly because each type of fibre requires a specialised neuron to fire them, and this can not be changed.
Gender or age differences?
It is believed there are no sex or age differences in fibre distribution, however, relative fibre types vary considerably from muscle to muscle and person to person. Sedentary men and women (as well as young children) have approximately 45% type 2 and 55% type 1 fibres.

Fibre types and sport
People at the higher end of any sport tend to demonstrate significant variation in patterns of fibre distribution from average people. For example, successful endurance athletes show a higher level of type 1 fibres.
Successful sprint athletes, on the other hand, have large numbers of type 2 b fibres in the key muscle groups.
Middle distance event athletes show approximately equal distribution of the 2 types. This is also often the case for power athletes such as throwers and jumpers.
Fibre adaptation to exercise
Endurance athletes

It has been well documented that elite endurance athletes possess a higher percentage of slow-twitch fibres in the muscles they use in their sport, compared to untrained individuals.
Is this due to genetic endowment or years of rigorous training? The answer is difficult to get at directly because we don't have comparative muscle biopsies of great athletes before and after they started training and excelling in their sport.
However, good basic investigation using experimental models has helped generate some answers. Unfortunately, almost all of this research suggests it is impossible to change the fibre type, without changing the motor neuron type also….in short, this is not going to happen!
So, why bother training so hard?
Skeletal muscles do respond to chronic overload (training), by trying to minimise the cellular disturbance caused by the training. Basically, this means they adapt to prevent too much exercise stress being put on the body.
With intense endurance training, fast-twitch fibres can develop more mitochondria and surrounding capillaries along with the slow fibres, so training improves the ability of your existing fibres to cope with the exercise stress you create for them.
Muscles are designed to produce force. Athletes train muscles to produce even more force in specific movements. A number of factors can have an impact on the ability of a muscle to produce force. Let's start with an obvious one.
The size of a muscle

Quite simply, a larger muscle has the potential to produce more force than a smaller muscle.
Keep in mind, a larger muscle doesn't always translate to better athletic performance, For example, throwing power, or a more powerful golf swing. A large set of biceps makes you better at bicep curls and filling out T-shirt sleeves! That said, if athletic form and function are well established, in some cases improving the size of key muscles will result in improved power output.
Injury to muscle or connective tissue

Damage to the muscle or connective tissue around a joint will lead to decreased force production of the muscles surrounding the joint. The brain is less likely to recruit musculature that causes pain. Trying to continue movement will most likely lead to movement compensation (for example, limping).
The number and type of motor unit recruited
Key Point
The larger the number of motor units recruited, the greater the force produced.
Fast-twitch fibres produce more force. Muscle size is achieved through developing primarily fast-twitch fibres. Those muscles that have a large proportion of fast-twitch fibres will be stronger than other muscles. This is the main factor contributing to muscle force generation.
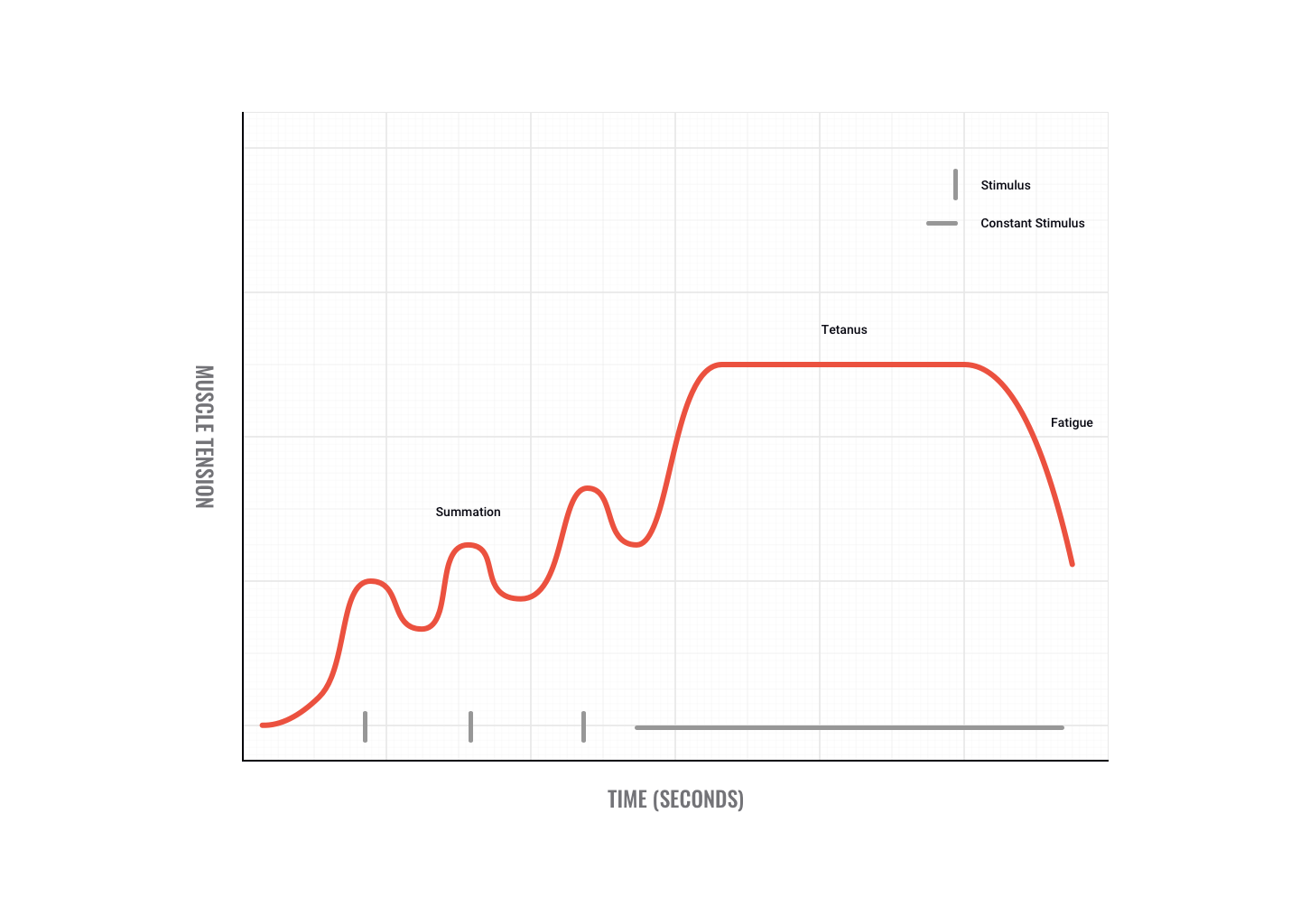
Tetanus
If motor impulses are sent fast and furious, twitches can sum to produce a smooth, sustained, maximum contraction called tetanus.
Maximum tetanic contractions produce a force approximately three to five times a single twitch contraction.
An example of when we use tetanus is when trying to overcome a heavyweight. Initially, we may not be able to move the weight but as we send more motor signals we recruit greater numbers of motor units and finally overcome the weight, e.g. pushing a car or performing a heavy dead-lift.
Stretch: length-tension relationship
Optimal length - Lo
- Maximum number of cross-bridges
- No overlap of actin fibres from opposite ends of the sarcomere
- Optimal working muscle range from 80-120% of Lo.
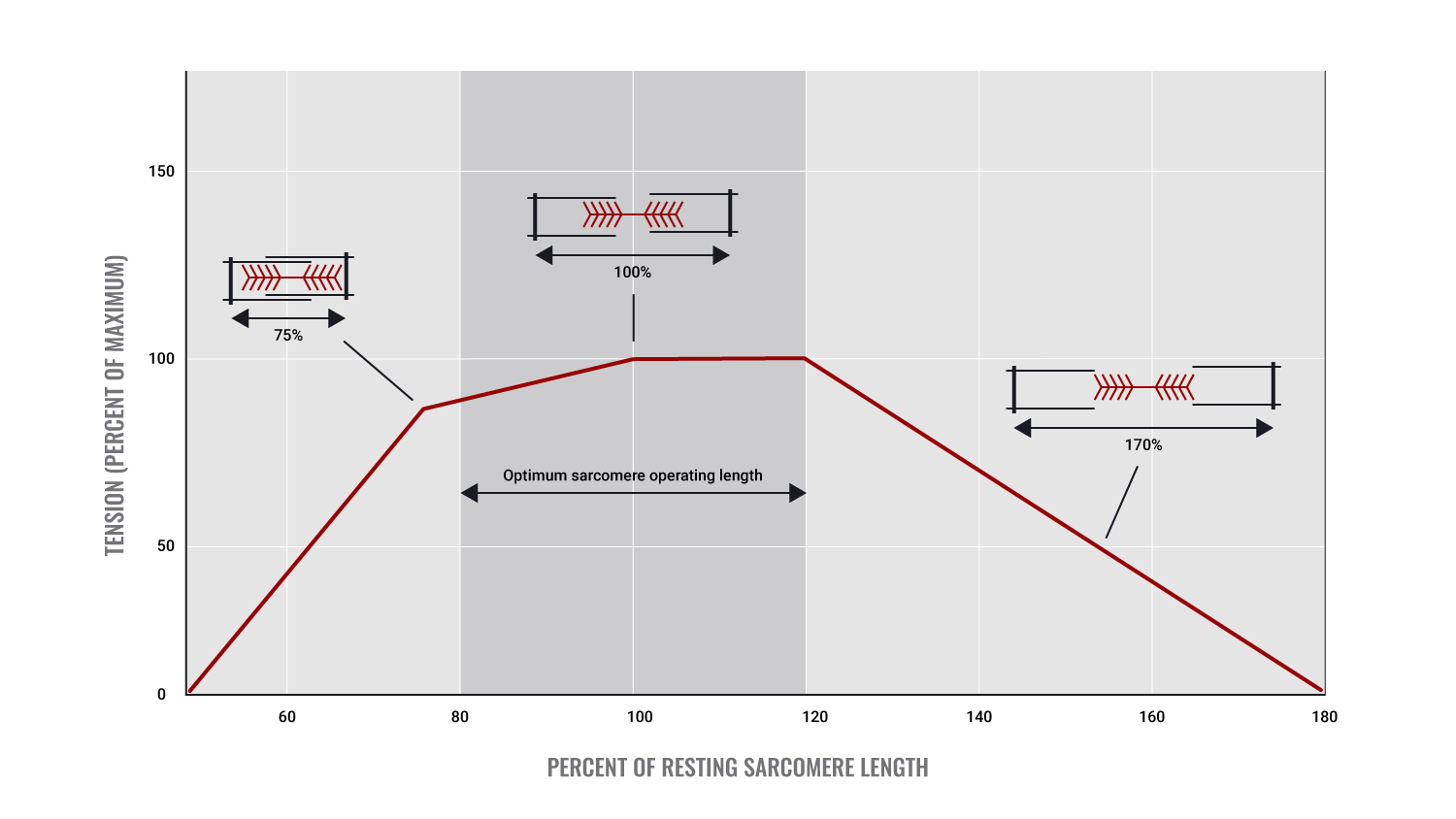
The starting length of a muscle prior to contraction
Starting length, more accurately referred to as "Resting length" is where each muscle has an optimal length for efficient/ optimal muscle contraction.
This length allows for maximal cross-bridge interaction and will lead to the greatest muscle tension being produced.
The rule governing this is known as the length-tension relationship (also known as the 'force-length relationship'). This states that if a muscle is either over-stretched or over-shortened prior to the initiation of contraction, muscle force will be reduced. For example, a 90° angle at the elbow joint is the ideal starting position for maximal force production by the biceps.
- Velocity of the Contraction
- Lighter loads can be moved faster
- Each muscle has it's own optimal speed of contraction
- If OPTIMAL velocity is achieved - force is MAXIMISED
- If the velocity of contraction is too fast - force is reduced as there is not enough time for all fibres to form cross-bridges - i.e. slow-twitch fibres can't fire fast enough to aid contraction
- Optimal velocity will allow for progressive recruitment of fibres. Therefore greater numbers of motor units are activated leading to greater force production.
Velocity and power
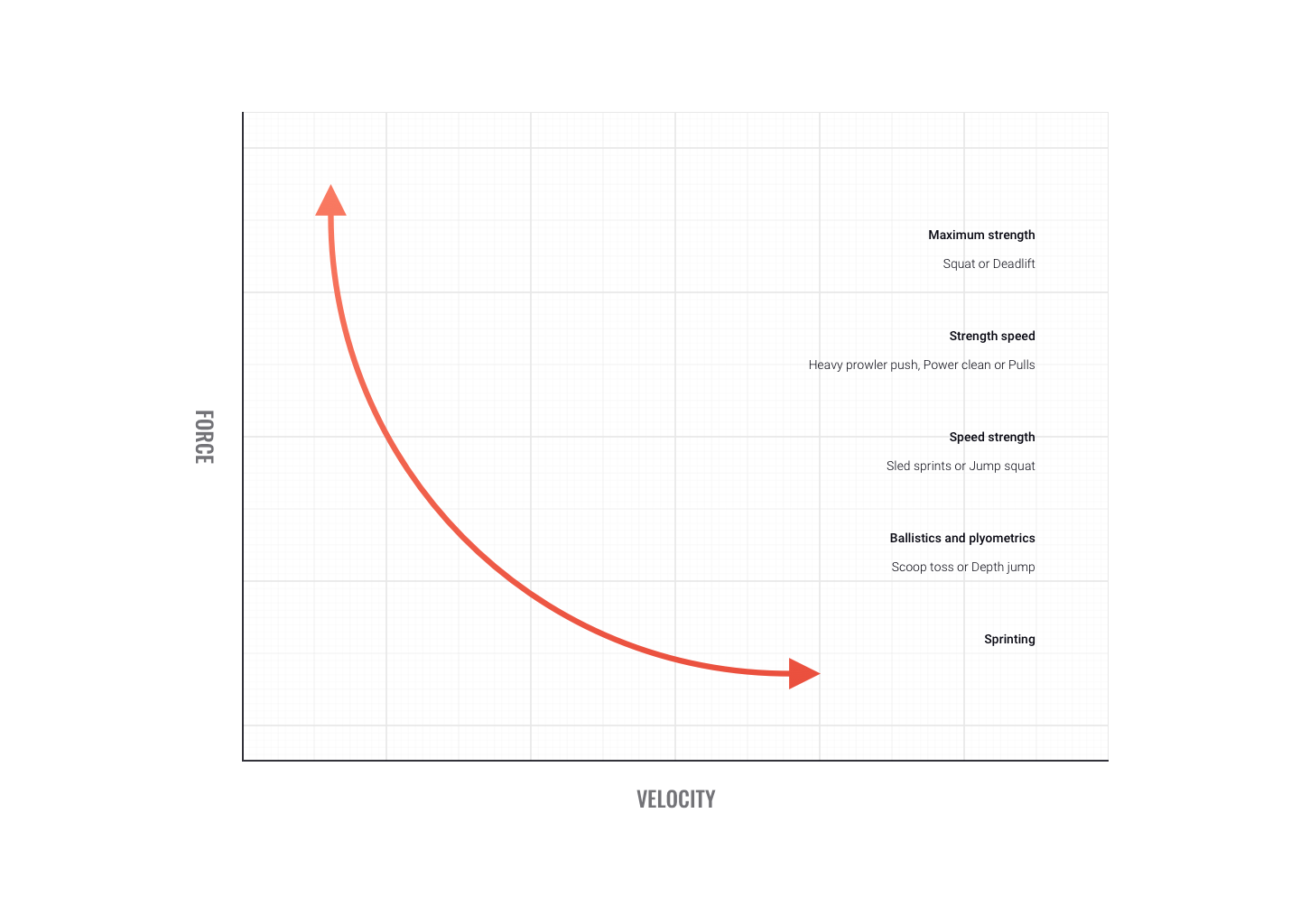
Some sports movements don't allow time for optimal force production. When force is produced quickly we term it 'power.
Power training targets fast-twitch fibres. While each muscle has it's own optimal velocity for power production, on average around 300 degrees/sec produces peak power. At velocities faster than this, power appears to plateau.
In this topic, we focused on the physiology of muscle and muscular contraction and the effects of exercise. You learnt about:
- muscular components and structure
- muscular contraction, including the:
- sliding filament theory
- all or none principle
- neuromuscular junction
- muscle fibre recruitment
- muscle fibre types
- genetics and muscle physiology
- muscular force, velocity and power, including:
- length-tension
- resting length
- muscular adaptation and exercise.
